Центральные эффекты СКЭНАР-стимуляции
К вопросу о функциональном назначении генерализованной
синхронизации медленноволновой активности структур мозга
Аппарат СКЭНАР Таганрогского ОКБ "РИТМ" применяется для лечения путем импульсной электрокожной стимуляции [З]. Актуальность исследования эффектов СКЭНАР-стимуляции определяется выявившимся в ходе лечебной практики необычайно широким диапазоном его оздоровительного действия [1,3,9,11]. Это предполагает, что СКЭНАР задействует некоторую общую систему организма, ответственную за его самовосстановление - систему "внутреннего доктора". Наш подход к этой проблеме был обоснован в предыдущей работе [14]. Мы предположили, что СКЭНАР активирует центральные механизмы управления самовосстановлением организма - прежде всего кору мозга и важнейшее звено вегетативной регуляции - гипоталамус при ведущей роли его переднего, парасимпатического отдела. Однако центральные эффекты действия СКЭНАР-стимуляции ранее не исследовались. Работа посвящена экспериментальному выяснению действия СКЭНАР-стимуляции на активность коры мозга и гипоталамуса млекопитающих.
Методика
Эксперименты проведены - на 8 обездвиженных тубокурарином морских свинках и на 2-х бодрствующих кроликах (12 опытов). Основной зоной для стимуляции СКЭНАР и на свинках, и на кроликах была поверхность плеча. Кожа в этой зоне очищалась от волос депеляторием. Для СКЭНАР-стимуляции использовали в основном коаксиальный электрод площадью 5 кв. см., и в отдельной серии испытаний - 24-х игольчатый электрод общей площадью около 2,5 кв. см. СКЭНАР-стимуляция осуществлялась на частоте 59,3 Гц. Силу стимуляции подбирали так, чтобы возникало локальное подрагивание кожи, не приводящее к попыткам избежать раздражения (в острых опытах - до проведения эксперимента). Во время такого раздражения животное вело себя спокойно.
В опытах на морских свинках обездвиженное тубокурарином животное переводилось на режим искусственного дыхания. Под действием кратковременного ветбуталового наркоза, соблюдая меры локальной анестезии, осуществляли необходимые операции для введения в мозг электродов. По выходе из наркоза для снижения болевого раздражения освобождали ушные проходы от держателей. Электроды вводили билатерально в структуры переднего и заднего гипоталамуса по стереотаксическим координатам атласа Tindal (1964). Корковые билатеральные отведения осуществляли эпидурально из зрительной, а также соматосенсорной коры, в зоне проекции плеча [15], - места приложения СКЭНАР-стимуляции. У кроликов в условиях ветбуталового наркоза и локальной анестезии - по стереотаксическим координатам И. П. Цветковой (1978) электроды вводили в структуры переднего и заднего гипоталамуса. Корковые отведения осуществляли через эпидуральные электроды билатерально - от соматосенсорной и зрительной коры. При проведении опыта кролик со свободноподвижной головой размещался в гамаке.
Результаты исследования и обсуждение
Для выяснения общих эффектов СКЭНАР-стимуляции в обычном режиме его использования, мы ориентировались на то, чтобы набрать достаточно большое число случаев его применения. С этой целью использовали относительно короткие - 2 мин. - серии стимуляции с частотой 59,3 Гц. Такая стимуляция применялась в 21 случае на свинках, и в 34 - на кроликах. По спектрам мощности сравнивали активность на 30 с участках записи перед стимуляцией, при действии раздражения через 1,5 мин. после начала стимуляции - чтобы дать эффекту развиться, и в последействии. На свинках и кроликах по спектрам мощности визуально отслеживались эффекты СКЭНАР-воздействия (рис. 1 - А, Б, В).
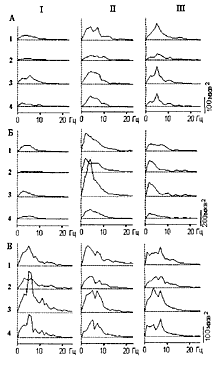
Рис.1 Примеры распределения частот в
спектрах мощности до (I), на фоне 2-х
минутной СКЭНАР-стимуляции (II), в
последействии (III) в структурах мозга
морской свинки (А, В), и кролика (Б).
(В) - пример «слома» доминирующего
ритма при СКЭНАР-стимуляции и
появление нового ритма, доминирующего
последействии
1 - зрительная кора;
2 - соматосенсорная кора;
3 - передний гипоталамус;
4 - задний гипоталамус
Эффекты преимущественно были связаны с ростом мощности в области медленноволновых колебаний в диапазоне альфа-дельта при большой разнице в рисунках конкретных спектров для различных отведении и для разных случаев стимуляции. Наблюдались перестройки ритмики (рис. 1.В). Анализ различных случаев свидетельствовал об увеличении мощности колебаний в области низких частот и о тенденции к развитию генерализованной активности по всем отведениям во время раздражения и после него.
На фоне вариаций прослеживалась общая тенденция к синхронизации медленноволновых колебаний уже при коротких, 2-х минутных, СКЭНАР-воздействиях. При этом в электрограммах наблюдали синхронизацию медленноволновой активности. Заметим, что подобную синхронизацию можно было наблюдать в ряде случаев сразу вслед за включением СКЭНАРа. Двухминутную СКЭНАР-стимуляцию терапевты в практике используют, применяя очень высокие - на грани терпимого - мощности стимуляции.
Для получения более отчетливых результатов мы использовали более длительную -30-минутную СКЭНАР-стимуляцию, такая длительность воздействия рекомендована для использования в практике СКЭНАР-терапевтов. При этом по-прежнему мы наблюдали и активность через 1,5 мин. после начала раздражения. Длительная стимуляция применялась для 4-х свинок, 12 опытов было проведено на бодрствующих кроликах. В этих случаях регулярно наблюдался рост медленноволновых колебаний и тенденция к их синхронизации в различных отделах мозга. У морских свинок при этом наблюдались альфа-подобные веретена (рис. 2-А). На электрограммах бодрствующих кроликов при длительной СКЭНАР-стимуляции кроме альфа-колебаний возникали вспышки колебаний в сигма-ритме и наблюдалось появление К-комплексов (рис. 2.Б).
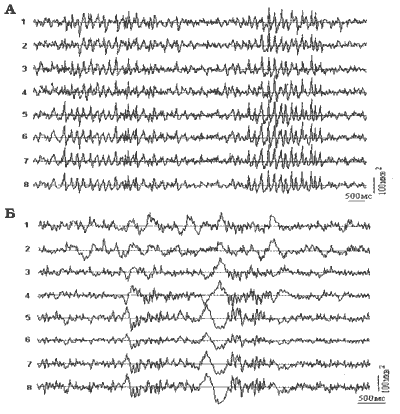
Рис.2 Примеры записи электрограммы структур мозга
морской свинки (А): 1,2 - зрительная кора; 3,4 - соматосенсорная кора;
5,6 - передний гипоталамус; 7,8 - задний гипоталамус
и кролика (Б): 1,2 - зрительная кора; 3,4 - соматосенсорная кора;
5,6 - задний гипоталамус; 7,8 - передний гипоталамус
Прослеживание эффекта СКЭНАР-воздействия по спектрам мощности при длительной стимуляции у кроликов показало моменты синхронизации активности в полосе не только альфаподобных колебаний, но и сигма-ритма - 13-15 Гц. Такие формы активности обнаруживаются во время сна, с его повышенным парасимпатическим тонусом [6,13]. Представляется, что их появление на фоне СКЭНАР-раздражения у кроликов также отражает рост парасимпатического тонуса.
В этой связи, мы особое внимание уделили наблюдениям за бодрствующими животными во время длительной СКЭНАР-стимуляции. Оказалось, что, несмотря на локальную кожную реакцию подрагивания, они перестают держать голову - кладут ее на гамак, ведут себя спокойно, глаза несколько западают - признаки характерные для повышения парасимпатического тонуса согласующиеся с фактом его активации. Известно [7], что при активации заднего гипоталамуса - повышении симпатического тонуса - характерно противоположное поведение животных: подъем головы, общее беспокойство, принюхивание. Сказанное выше указывает на то, что хотя во время генерализованной синхронизации активности изменяются амплитуды медленных волн и в переднем, и в заднем гипоталамусе (рис.1, 3), эти изменения можно рассматривать, как происходящие при ведущей роли переднего, парасимпатического, гипоталамуса.
При прослеживании динамики развития эффекта СКЭНАР-стимуляции по спектрам мощности рост медленноволновых колебаний во всех исследуемых отведениях отмечался регулярно. В отдельных случаях максимальная синхронизация наблюдалась уже через полторы минуты (короткая стимуляция) и сразу вслед за началом воздействия (рис. 3-Б), но чаще она развивалась на протяжении первых 15 мин. стимуляции (рис. 3-А,В.). Повторяемость появления генерализованной синхронизации от опыта к опыту свидетельствует о закономерности этого события (р < 0,01, критерий знаков) как результата СКЭНАР-воздействия.
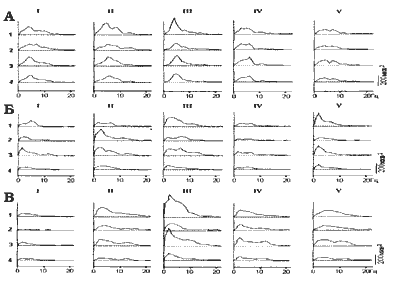
Рис.3 Примеры динамики развития активности в структурах мозга при
длительной СКЭНАР-стимуляции, прослеживаемой по спектрам мощности
для морской свинки (А) и кролика (Б) - плечо, (В) - голова в фоне (I),
через 1,5 мин. стимуляции (II), через 15 мин. стимуляции (III), через З0 мин.
стимуляции (IV) и в последействии (V) 1 - зрительная кора;
2 - соматосенсорная кора; 3 - передний гипоталамус; 4 - задний гипоталамус.
Чтобы получить общее представление об эффективности короткой и длительной СКЭНАР-стимуляции и выяснить особенности действия СКЭНАРа на различные области мозга, сопоставили усредненные спектры мощности, полученные для случаев короткой СКЭНАР-стимуляции с суммарными спектрами, характеризующими моменты максимальной синхронизации при длительной стимуляции (рис.4-А, Б).
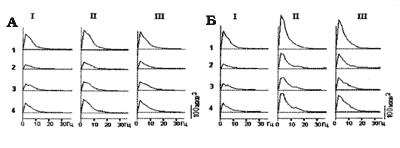
Рис. 4 Сравнение суммарных картин активности мозга кролика для
короткой (А) и длительной (Б) СКЭНАР-стимуляции. I - до, II - на фоне
СКЭНАР-стимуляции, III - в последействии. 1 - зрительная кора;
2 - соматосенсорная кора; 3 -задний гипоталамус; 4 - передний гипоталамус.
Из рисунка видно, что тенденция к общему росту мощности медленных колебаний сравнительно с предшествующим фоном, обнаружившаяся при короткой стимуляции, при длительной стимуляции усиливалась, развиваясь в отчетливый эффект (рис. 4-Б). Синхронизация была лучше выражена для переднего гипоталамуса, чем для заднего и для зрительной коры, чем для соматосенсорной. Такая картина сохранялась в последействии. Факт более выраженной активности переднего гипоталамуса по сравнению с задним свидетельствует о ведущей роли первого и указывает на повышение парасимпатического тонуса под действием СКЭНАР-стимуляции. По критерию знаков рассмотрели согласованность изменений активности в зрительной коре и переднем гипоталамусе. Оказалось, что активность этих структур высоко скоррелирована - Р < 0,01 и для кроликов и для свинок - т. е. в целом по выборке Р < 0,001. Это обстоятельство позволяет судить о повышении активности переднего гипоталамуса (рост парасимпатического тонуса) по показателям синхронизированной активности зрительной коры.
Факт большей реакции зрительной коры, чем соматосенсорной на электрокожное раздражение требует особого рассмотрения. Надо учесть, что наблюдаем мы уже развившийся системный эффект воздействия, который захватывает многие структуры мозга. Видимо, очаг активности, создаваемый СКЭНАР-стимуляцией изначально возникает в соматосенсорной коре. Это вызывает активацию синхронизирующей системы с ее важным звеном - передним гипоталамусом. Его активация, как известно, сопровождается повышением парасимпатического тонуса. В свою очередь передний гипоталамус, как это следует из наших результатов и как это было подробно рассмотрено Н. В. Сунцовой [13], вызывает генерализованную синхронизацию, характеризующуюся высокой скоррелированностью в области альфа-подобных колебаний для переднего гипоталамуса и зрительной, но не соматосенсорной коры. Повышенная активность переднего гипоталамуса приводит к повышенной альфа-активности и в последействии. В развитии синхронизированной активности возможна особая роль медленнопроводящих волокон спиноталамического тракта, часть которых оканчивается в гипоталамусе. Предложенная, пусть неполная схема взаимодействии, позволяет экспериментальную проверку.
Заслуживает внимание, что СКЭНАР-воздействие часто характеризуется относительно высокой амплитудой альфа-подобной активности в зрительной коре. Согласно исследованиям Гаркави с сотрудниками [2] такая электрофизиологическая картина есть коррелят антистрессорной реакции тренировки - у животных и человека- С ней связывают важные оздоровительные последствия. Именно с развитием антисрессорных реакций эти исследователи связывают лечебный эффект аппарата СКЭНАР. Наши результаты показывают, что развитие и сохранение в последействии повышенной альфа-подобной активности в зрительной коре(рис 1.А,Б), особенно, после длительной стимуляции, (рис, 3.В) может рассматриваться и как критерий эффективности СКЭНАР-воздействия, и как указание на парасимпатическую природу этих реакций. В свете приведенных данных особый интерес представляют отставленные эффекты последействия, о чем несколько ниже.
Известно, что стимуляция кожи черепа, характеризующаяся преимущественно парасимпатической иннервацией [8] имеет особо хорошие лечебные последствия. Мы исследовали эффекты стимуляции кожи головы (рис. З.Б,В) - раздражение осуществлялось через 24-х игольчатый электрод. В согласии с отмеченным выше клиническим эффектом, результаты такой СКЭНАР-стимуляции оказались более выраженными. Эффект СКЭНАР-стимуляции кожи черепа характеризуется большей синхронизацией активности, чем при стимуляции кожи плеча через тот же, игольчатый электрод (рис. 5). Интересно отметить, что при стимуляции кожи черепа поведение кролика стало похожим на сон - он закрыл глаза. Это указывает на значительный рост парасимпатического тонуса. У этого кролика в последействии через сутки наблюдали повышенную медленноволновую активность.
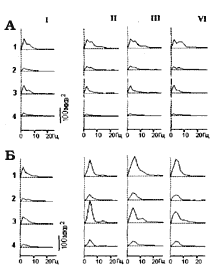
Рис. 5 Примеры последействия
фоновой активности структур мозга
кролика через сутки после
длительной СКЭНАР-стимуляции
плеча (А) и головы (Б).
1 - зрительная кора;
2 - соматосенсорная кора;
3 - передний гипоталамус;
4 - задний гипоталамус;
I - до, II, III, IV - после с интервалом 15 мин.
Отмеченные выше моменты синхронизированной медленноволновой активности, как можно заключить на базе полученных данных, представляют собой основной электрофизиологический коррелят эффективного действия СКЭНАР. В целом состояние мозга при этом похоже на переходное ко сну. В этой связи заслуживает внимания факт, что седативное действие с переходом ко сноподобному состоянию отмечается и как общий эффект при иглотерапии [обзор 5] - в том числе и у животных. Это совпадение, видимо, не случайно, так как между отмеченными средствами лечения при многих различиях имеется и существенное сходство.
Роль моментов синхронизации медленноволновой активности, создаваемых СКЭНАРом, можно усмотреть в том, что они облегчают доступ информации о "внутреннем теле" организма к экрану коры;, хранящему в себе эталон здорового тела. В условиях активного бодрствования, характеризующегося высоким симпатическим тонусом, как известно, кора мозга доступна лишь очень мощным сигналам от внутренних органов. Иное дело во сне с его высоко синхронизированной активностью [4,6,10].
Представляется, что аналогично тому во сне генерализованная синхронизация, создаваемая при СКЭНАР-воздействиях, обеспечивает возможность доступа сигналов о состоянии "внутреннего тела" организма к коре и выработку соответствующих корректирующих сигналов к промежуточным уровням управления и исполнительным структурам. - в первую очередь - к структурам переднего гипоталамуса, перестраивающего работу вегетативной системы на режим восстановления. При этом благодаря фокусу активности, создаваемому СКЭНАРом, указывается локализация очага поражения. Такое состояние не сон, при котором генерализованная активность весьма регулярна. Но о сходстве состояний свидетельствует сонливость, развивающаяся по наблюдениям клиницистов при эффективном действии СКЭНАРа. Самонаблюдения после СКЭНАР-стимуляции также подтверждают этот эффект.
Общая схема действия СКЭНАРа в свете полученных данных, показывающих в числе прочего и рост синхронизированной активности в соматосенсорной коре, представляется следующим образом. Очаги активности, создаваемые СКЭНАР-стимуляцией, в соматосенсорной коре 1 и 2, через лемнисковую и экстралемнисковую системы, а также, возможно, и прямые влияния на гипоталамус по спиноталамическому тракту активирует синхронизирующую систему мозга с передним, парасимпатическим гипоталамусом. Тот, в свою очередь, поддерживает общую синхронизацию коры и подкорковых структур, в том числе и особенно - в диапазоне альфа-ритма в зрительной коре. В этих условиях отделы коры мозга, хранящие в себе эталон здорового "внутреннего тела", становятся доступны для сигналов от внутренних органов с очагом повреждения, указываемым в связи с локализацией СКЭНАР-электрода. Становится возможной выработка корректирующих сигналов на управляющие и исполнительные системы организма, гипоталамуса прежде всего, работа которых координируется на самовосстановление. В последействии повышенная активность переднего гипоталамуса обеспечивает высокую амплитуду альфа-ритма в зрительной коре. Т. е., при достаточном парасимпатическом тонусе, отражающем деятельность организма по самовосстановлению должно наблюдаться повышение мощности альфа-активности в затылочных отведениях. Именно это и обнаруживается в последействии СКЭНАР-стимуляции.
Выводы
1. Обнаружен центральный эффект СКЭНАР-стимуляции, заключающийся в генерализованной активности коры мозга и стркуктур гипоталамуса среди них ведущую роль играет активация переднего гипоталамуса, сопровождающаяся повышением парасимпатического тонуса.
2. Подтверждена высокая скорелированность активации переднего гипоталамуса с повышением альфа-подобной активности затылочных отведении. - зрительная кора мозга. Т.о., повышение альфа-активности в зрительной коре может служить показателем повышения активности переднего гипоталамуса и соответственно парасимпатического тонуса.
3. Эффект СКЭНАР-стимуляции сохраняется в последействии и может обнаруживаться спустя сутки после проведения стимуляции.
Список литературы
1. Гаркави Л.Х. и др. Некоторые биофизические подходы к механизму действия аппарата СКЭНАР // В сб.: СКЭНАР-терапия, СКЭНАР-экспертиза.
Таганрог: 1997. Вып. 3. с. 13.
2. Гаркави Л.Х. и др. Антистрессорные реакции и активационная терапия.
М.: ИМЕДИС. 1998. 656 с.
3. Гринберг Я.3. Эффективность СКЭНАР-терапии. Физиологические аспекты. // В
сб.: СКЭНАР-терапия, СКЭНАР-экспертиза. Таганрог. 1998 Вып. 4. с. 8.
4. Данилова Н. Н. Психофизиологическая диагностика функциональных состояний.
М. 1992. 192 с.
5. Игнатов Ю.Д. и др. Акупунктурная анальгезия. Л. 1990. 256 с.
6. Карманова И.Г. Что мы знаем о цикле "бодрствование-сон" и причинах его
нарушения. СПб. 1998. 93 с.
7. Козловская М.П., Вальдман А.В. Фармакология ретикулярной формации и
синаптической передачи. М. 1963. 217 с.
8. Лувсан Гаваа. Традиционные и современные аспекты восточной медицины.
М. 1992. 576 с.
9. Ляшедько П.П. СКЭНАР-терапия в лечении сочетанной черепно-мозговой
травмы. // В сб.: СКЭНАР-терапия, СКЭНАР-экспертиза. Таганрог. 2000. Вып.6. с.27.
10. Ноздрачев А.Д. и др. Общий курс физиологии человека и животных. // Кн. 2.
Физиология висцеральных систем. М.: Высш. шк. 1991. 528 с.
11. Окишева Т.А. Случай лечения нейросенсорной тугоухости.
// В сб.: СКЭНАР-терапия, СКЭНАР-экспертиза. Вып. 6. с. 101.
12. Самохин А.В., Готовский Ю.В. Электропунктурная диагностика по методу Р.
Фолля. М.: ИМЕДИС. 1993. 214 с.
13. Сунцова Н.В. Переднемозговые механизмы развития сна.
// Дисс. на соиск. уч. степени д. б. н. - РГУ, Ростов на Дону, 2000. 372 с.
14. Чебкасов С.А. Стратегия здоровья. Система опережающего самовосстановления
биоструктур. Проблема активации парасимпатической вегетативной системы.
Валеология. 2000. № 1. с. 80.
15. Zeigler H.P. Cortical sensory and motor areas of the guinea pig.
// Arch. Ital. Biol, 1964. V. 162, fasc. 4. P. 587.
С. А. Чебкасов, Ю. И. Берешполова
|
Научно-исследовательский институт нейрокибернетики им. А.Б. Когана |




